Principi
di Biologia Cellulare
PIASTRINOPOIESI
Le piastrine sono piccole cellule enucleate, altamente specializzate, con un
volume medio di 7 fl. e originate dai megacariociti.
Il loro numero nel sangue circolante è, normalmente, compreso tra 150.000 e
400.000/µl; l'emivita è di 5-8 giorni.
Le piastrine presentano una forma ed un volume piuttosto eterogenei.
I cambiamenti di forma dipendono dalle differenti attività: mentre allo stato
di "riposo" sono lenticolari, durante il processo di attivazione assumono una
forma sferica irregolare che presenta lunghi e sottili pseudopodi.
Il volume dipende dall'età delle piastrine: le più giovani presentano un volume
maggiore rispetto a quelle vecchie e contengono una maggiore quantità di RNA.
Un'altra caratteristica che permette di identificare l'età delle cellula è rappresentata
dalla densità piastrinica, definita dal contenuto in granuli citoplasmatici:
quando è minore si tratta di una cellula matura.
Le piastrine circolanti vengono liberate dai megacariociti maturi a livello
midollare e, in piccola parte, anche a livello polmonare.
Le piastrine rilasciate nei sinusoidi midollari ed immesse in circolo si distribuiscono
in un pool circolante vero e proprio (circa 70%) e in un pool splenico (circa
30%).
Le piastrine presenti nel sangue circolante in piccola parte vengono consumate
durante il processo emostatico ed in maggioranza muoiono per senescenza.
La piastrinopoiesi è pienamente efficace, quando, ad una data massa megacariocitaria
corrisponde una produzione piastrinica ottimale.
E' importante ricordare che non esiste un pool midollare piastrinico di riserva
e che il tempo di transito midollare dei megacariociti non appare passibile
di modificazioni significative.
STRUTTURA PIASTRINICA
Nella figura 2.1 viene illustrata una piastrina allo stato di "riposo".
Come si può osservare la piastrina è racchiusa da una membrana trilaminare che
porta, sulla sua superficie esterna, fibrinogeno e molti altri fattori plasmatici
della coagulazione.
Questa membrana citoplasmatica presenta una struttura a doppio strato lipidico
in cui si trovano immerse glicoproteine (complessi di carboidrati e proteine)
che hanno una fondamentale importanza nel processo emostatico per il loro ruolo
di recettori o di proteine adesive; in effetti mediano le più importanti funzioni
delle piastrine che sono l'adesione, l'aggregazione e il trasporto delle molecole.
La forma discoidale della piastrina è mantenuta dal citoscheletro, che è formato
dal sistema microtubulare e da quello contrattile costituiti principalmente
da proteine quali l'actina, la miosina e la trombostenina.
A atmosfera
piastrinica
B membrana trilaminare
C sistema canalicolare aperto
D citoscheletro (sistema microtubulare contrattile )
E sistema tubulare denso
F glicogeno
G mitocondrio
H granuli α ( b-TG , FP4, fibrinogeno, VIII-Ag )
I granuli δ ( ADP, ATP, serotonina, Ca++ )
L granuli λ ( idrolisi, proteasi, catepsine ) |
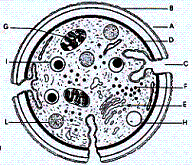 |
| Figura
2.1 - Struttura di una piastrina a "riposo" vista in sezione ( Sante Tura). |
Nel citoplasma
sono presenti alcuni mitocondri e specifiche strutture vescicolari quali: i
granuli a (densi), i granuli ß e i granuli λ (lisosomi).
Nella tabella 2.1 vengono riassunte le caratteristiche dei singoli granuli contenuti
all'interno delle piastrine.
| Granuli |
Diametro |
Contenuto |
| α-granuli |
0,15-0,40 |
PF4,β-tromboglobulina,
fibronectina, vitronectina, trombospondina,fibrinogeno,vWF, FV, FVIII, FXIII-a,
inibitori del t-PAPDGF, EGF,TGF-β, IGF-I, IGF-II, HGF |
| ó-granuli
(corpi densi) |
0,17 |
ADP, ATP,
GTP. GDP, fosfato, ortofosfato, calcio-ioni,magnesio-ioni, serotonina, adrenalina,istamina |
| λ-granuli
(lisosomi) |
0,17-0,25 |
fosfatasi
acida, aril-solfatasi, Betaglucoronidasi, galattosidasi |
|
Tabella
2.1. - Caratteristiche
dei granuli piastrinici.
|
FUNZIONE PIASTRINICA
La funzione delle piastrine è quella di formare un tappo emostatico, dove c'è stata una lesione dell'endotelio, per impedire la fuoriuscita di sangue (figura
2.2)2.
Le piastrine circolano senza aderire quando il vaso è intatto e ciò avviene,
probabilmente, per effetto di vari fattori, quali le cariche di membrana, la
presenza di mucopolisaccaridi simili all'eparina, la presenza di inibitori delle
proteasi e la formazione di prostaciclina da parte delle cellule endoteliali.
Di contro, quando il vaso subisce una lesione, le piastrine aderiscono, stimolate
dall'esposizione delle fibre collagene e della membrana basale ed influenzate
dalla presenza della componenente von Willebrand del fattore VIII e della fibronectina.
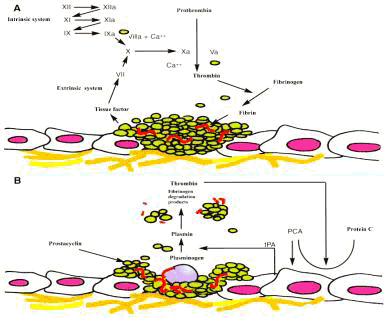 |
Figura 2.2.
- Rappresentazione schematica della fase vascolare (A) e piastrinica(B)
(Madon, Gabutti, Miniero, 1998) |
All'adesione segue l'aggregazione, una reazione totalmente dipendente dalla
capacità delle glicoproteine IIb-IIIa di legare il fibrinogeno plasmatico, che
avviene seguendo delle fasi ben precise1 (figura 2.3).
Innanzitutto, si verifica un cambiamento di forma perché durante questa fase
la piastrina reagisce allo stimolo aggregante modificandosi da discoidale in
sfera spinosa; successivamente, per effetto dell'agente aggregante esogeno (tabella
2.2) le piastrine tendono ad avvicinarsi l'una all'altra e centralizzano i
granuli intracellulari; tuttavia, se lo stimolo non è sufficiente, gli aggregati
si disperdono, i granuli si ridistribuiscono all'interno del citoplasma e la
piastrina ritorna allo stato di riposo (aggregazione primaria: fase reversibile).
Di contro, se lo stimolo aggregante esogeno è stato sufficiente, si attiva il
metabolismo dell'acido arachidonico ed i granuli centralizzati liberano il loro
contenuto (reazione di liberazione); infine, le sostanze contenute nei granuli
(ADP, serotonina ecc.) e quelle formate dal metabolismo dell'acido arachidonico
(prostaglandine e trombossani) aggregano le piastrine irreversibilmente (aggregazione
secondaria: irreversibile).
 |
Figura 2.3 - La figura illustra i cambiamenti di forma e la perdita dei
granuli da parte delle piastrine che vanno incontro ad aggregazione irreversibile
(Sante Tura, 1997). |
| Platelets |
| Principal
Aggregant Agents |
| Physiological
agents |
collagen |
Thrombin
ADP Arachidonic Acid Adrenalin, Noradrenalin Serotonin |
| Non physiological
agents |
ristocetin |
Bovin fibrinogen
Ca++ ionophores |
Tabella 2.2 (Sante Tura, 1997). |
Il meccanismo biochimico responsabile del fenomeno di aggregazione piastrinica è l'aumento del livello intracitoplasmatico degli ioni Ca++, evento regolato
dall'AMP ciclico e dal metabolismo dell'acido arachidonico.
L'aggregazione piastrinica è un fenomeno molto studiato ma non ancora completamente
conosciuto.
L'interesse che caratterizza questo problema è legato al risvolto clinico e
terapeutico.
Sulla base delle attuali conoscenze, si può ipotizzare che esistano tre vie
di aggregazione piastrinica: la prima è mediata dall'ADP esogeno ed endogeno;
la seconda è mediata dall'acido arachidonico; la terza, anche se non completamente
chiarita, è mediata dal PAF (Platelet Activating Factor), una lisolecitina liberata
da piastrine attivate, macrofagi, neutrofili, basofili e probabilmente da altre
cellule.
L'acido arachidonico e il PAF possono aggregare le piastrine direttamente, oppure
mediante la liberazione di ADP.
Una volta che si è formato l'aggregato, i prolungamenti citoplasmatici delle
piastrine si annodano ai tralci di fibrina in modo che il trombo possa consolidarsi.
Infine, le piastrine, con un meccanismo non chiarito, grazie alle proteine contrattili
del citoscheletro, si retraggono.
Le piastrine, durante la reazione di liberazione, mettono a disposizione del
processo emocoagulativo il fattore piastrinico 3 o FP3, importante per la via
intrinseca della coagulazione plasmatica1.
Nella figura 2.4 viene schematizzato il ruolo svolto dalle piastrine nella formazione
del trombo adeso alla parete vascolare.
Una volta che le piastrine sono state stimolate ad aderire alla parete vascolare,
le sostanze in esse contenute vengono liberate spontaneamente.
A loro volta queste sostanze promuovono l'aggregazione di nuove piastrine, che è favorita dal rilascio del fattore di Von Willebrand; questa sostanza è adesiva
per la proteina di membrana GP1b e per il fibrinogeno.
Le piastrine attivate liberano anche ADP e trombossano A2, i quali richiamano
ulteriori piastrine, che subiscono alterazioni di forma e liberano granuli,
causando ulteriore aggregazione.
Le proteine piastriniche di membrana GP2b e GP3a aderiscono alla fibrina e al
fibrinogeno in un processo che tende a stabilizzare il trombo che si sta formando.
Figura 2.4.
Ruolo delle piastrine nella formazione del trombo (modificato da Rubin e
Faber, 1991 ). |
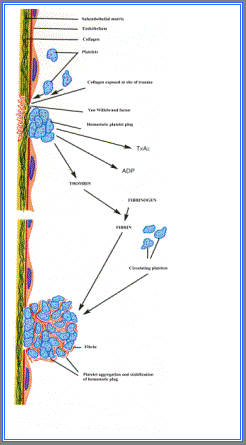 |
Il fattore più importante per la progressione e la stabilizzazione del trombo
durante la coagulazione è, verosimilmente, la produzione di trombina, che viene
prodotta nell'area danneggiata sia dalla via intrinseca che dalla via estrinseca
(figura 2.5).
La via estrinseca inizia con il rilascio di tromboplastina da parte delle cellule
danneggiate, mentre la via intrinseca prende avvio dall'attivazione del fattore
XII (fattore di Hageman), che dipende dal legame di questo fattore con un componente
delle cellule danneggiate.
La trombina stessa è sufficiente per stimolare l'ulteriore rilascio di granuli
piastrinici e il susseguente richiamo di nuove piastrine.
A mano a mano che la coagulazione procede, per azione della trombina sul fibrinogeno,
si forma la fibrina che crea ponti crociati con i recettori del GP2b e del GP3a
delle piastrine.
Questi legami stabilizzano l'aggregazione delle piastrine e la loro adesione
alla superficie sottostante.
Infine le piastrine, con un meccanismo non chiarito, per opera delle proteine
contrattili del citoscheletro, si retraggono.
FIBRINOLISI
Affinché venga mantenuto l'equilibrio della bilancia emostatica-emocoagulatoria,
è necessario ristabilire il flusso ematico; pertanto, sono indispensabili sistemi
di controregolazione che fisiologicamente modulino ed inibiscano il processo
emocoagulatorio.
Il sistema della fibrinolisi (figura 2.6) presenta strette analogie con il
sistema della coagulazione: in condizioni fisiologiche entrambi si trovano in
uno stato di equilibrio dinamico, in quanto regolati da attivatori ed inibitori.
La fibrinolisi ha varie funzioni:
1) degradare i complessi solubili di fibrina;
2) limitare la formazione del tappo emostatico nelle sedi di danno vascolare;
3) rimuovere la fibrina al termine dei processi riparativi.
L'attività fibrinolitica ha un ritmo circadiano: bassa al mattino, aumenta nel
corso della giornata, riducendosi poi nella notte.
La molecola centrale del sistema della fibrinolisi è costituita dal plasminogeno,
proenzima inattivo che venendo convertito nella forma attiva plasmina diventa
in grado di lisare la fibrina generando dei prodotti di degradazione solubili
che sono normalmente rimossi dal circolo ad opera del sistema macrofagico (figura
2.6).
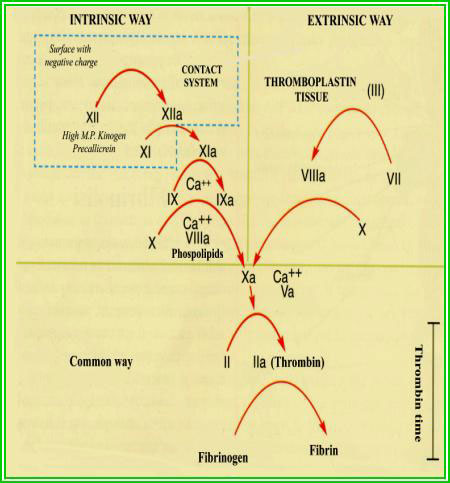 |
|
Figura 2.5.
- Cascata enzimatica della coagulazione.
|
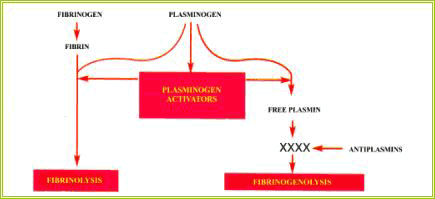 |
| Figura 2.6.
- Schema della regolazione del processo fibrinolitico. |
GRANULI
PIASTRINICI
I granuli sono l'organulo più rappresentato nel citoplasma delle piastrine e
sono visibili al microscopio ottico anche se è difficile apprezzarne l'individualità e la differenza dai mitocondri.
I granuli sono stati inizialmente designati con lettere dell'alfabeto greco,
ma attualmente solo il termine granuli a α è in uso.
I cosidetti granulomeri ß sono in realtà mitocondri mentre i granulomeri γ sono
spazi chiari senza struttura definita, come vacuoli e vescicole, che con tutta
probabilità fanno parte del sistema canalicolare.
I granulomeri δ, considerati molto rari, sarebbero dei siderosomi, mentre si
definivano granulomeri ε i depositi di glicogeno.
Ai fini dei processi di guarigione e della rigenerazione ossea, l'attenzione
deve essere rivolta ai granuli piastrinici a α che contengono oltre a proteine
plasmatiche e fattori della coagulazione, quei fattori di crescita che si ritiene
essere i mediatori della guarigione e della rigenerazione.
E' stato dimostrato che tre fattori di crescita sarebbero importanti sia nel
processo di guarigione che di rigenerazione: il PDGF, il TGF-ß e, recentemente,
sono stati scoperti anche l'IGF-I e II.
Questi fattori si possono definire "gli ormoni della guarigione" delle ferite
e, pertanto, ogni volta che si crea una ferita il nostro organismo per un meccanismo
di sopravvivenza mette in atto, immediatamente, dei meccanismi di guarigione
che possono essere di riparazione o di rigenerazione.
La vita media di una piastrina in una ferita e la diretta influenza dei fattori
di crescita rilasciati dai granuli a in essi contenuta, è di circa 5 giorni.
Pertanto, l'estensione della guarigione e l'attività di rigenerazione ossea
sono determinati da 2 meccanismi:
1) incremento ed attivazione delle cellule staminali midollari a differenziarsi
in osteoblasti che possono così secernere TGF-ß e IGF che a loro volta stimolano
la deposizione di matrice osteoide;
2) chemiotassi dei macrofagi, che venendo richiamati nell'innesto osseo, possono
sostituire le piastrine come fonte primaria di fattori di crescita a partire
dal terzo giorno.
I macrofagi vengono attratti verso l'innesto sia dalla liberazione di PDGF dai
granuli piastrinici che dalla presenza di un gradiente di ossigeno superiore
a 20 mmHg che deve necessariamente essere presente tra lo spazio morto dell'innesto
(pO2 = 5-10 mmHg) e il tessuto adiacente che presenta una tensione di ossigeno
normale (pO2 = 45-55 mmHg).
Si ritiene, quindi, che i fattori di crescita derivanti dal macrofago possano
essere identici al PDGF rilasciato dalle piastrine, così che le cellule staminali
midollari vengono indotte a secernere ulteriormente TGF-ß e IGF-I e II in modo
da continuare la stimolazione della formazione di osso, quale risposta autocrina.
BIBLIOGRAFIA
Sante Tura (ed): Lezioni di Ematologia. Società Editrice ESCULAPIO, Bologna,
1997.
Madon E., Gabutti V., Miniero R. (eds): Ematologia e Oncoematologia Pediatrica.
McGraw-Hill libri Italia srl, Milano, 1998.
Rubin Emanuel., Faber John L. (eds.): Patologia Generale- Atlante. McGraw-Hill
libri Iralia srl, 1991.
Beutler E, Lichtman MA, Coller BS, Kipps T (eds):.Williams Heatology, Fifth
Edition, McGraw-Hill, Inc., 1995.
Lynch SE:, Genco RJ, Marx RE (eds): Tissue Engineering: Applications in Maxillofacial
Surgery and Periodontics. Chicago: Quintessence; 1999.